GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By David B. Wahl & Ian D. Gauld
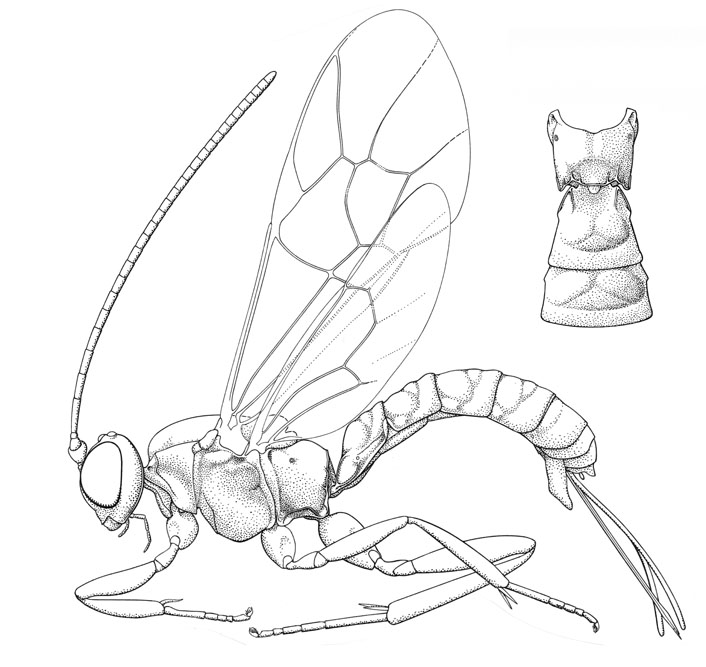
Lycorina scitula |
The subfamily Lycorininae is an enigmatic, small group of moderate-sized ichneumonids placed in a singe genus, Lycorina. Most are strikingly brightly coloured, handsome insects that are easily recognized by the metapleural process and the impressed triangular pattern on the metasomal tergites. Much remains to be discovered about the biology of these cryptic insects, but all recorded hosts are the larvae of small species of Lepidoptera.
Classification and diversity
Lycorinines may easily be distinguished from all other ichneumonids as they possess two quite conspicuous autapomorphic features:
- T1-4 each with a central triangular area bounded by deep impressions.
- presence of a metathoracic hook that extends backwards to engage a protuberance on the propodeum.
The Lycorininae contains a single genus, Lycorina. Classical authors put the genus in the Pimplinae. Townes, in his various reclassifications of ichneumonids, initially placed it as a tribe in the Banchinae (Townes & Townes, 1951), later as a genus in the banchine tribe Glyptini (Townes et al., 1965) and finally, on the basis of the mature larva, as a separate subfamily (Townes, 1970). The Lycorininae is a taxonomically rather isolated group, with no obviously close affinities to other lchneumonidae. but the characters supporting this association are plesiomorphies, such as possession of an unspecialized T1 with the spiracles positioned before the center. Lycorinines superficially resemble some glyptines, such as Zaglyptomorpha species, and both have grooves on the metasomal tergites, an epomial crest and an anteriorly expanded submetapleural carina. These similarities, however, seem to be superficial, as the form and position of the metasomal grooves differ, and lycorinines possess a distinctly impressed subocular groove and do not have a notched ovipositor.
Lycorina is a small cosmopolitan genus consisting of 28 described species, with a few species in almost all parts of the World (Yu & Horstmann, 1997). Several fairly distinctive species-groups are recognizable - and some of these have, in the past, been accorded generic distinction. Townes (1970) recognized three genera: Gonioglyphus, Lycorina, and Toxophoroides. Interrgrades occur between some of these groups, and there are other quite distinctive species groups which have never been accorded generic status, leading Gauld (1984) to treat all species as belonging to a single genus, Lycorina.
The general biology of Lycorininae
The focal host range appears to be weakly concealed larvae of microlepidoptera. Some northern temperate species have been reared from Crambidae in leaf-rolls or webs (Doerksen & Neunzig, 1974; Finlayson, 1976) and Tortricidae (Chao, 1980; Shaw, 2004). In Costa Rica Lycorina apicalis has been reared as a parasitoid of a species of Lamprosema (Lepidoptera: Crambidae) (Gauld, 1997) and from the prepupae of Ethmia catapeltica Meyrick (Ethmiidae) (Coronado-Rivera et al., 2004).
Until recently, the only evidence for ecto- vs. endoparasitism in lycorinines came from larval morphology. The cephalic capsule of the final instar larva (Finlayson, 1976; Short, 1978; Chao, 1980) possesses features of both ecto- and endoparasitic ichneumonids. The mandibular teeth of the final instar larva are denticulate, resembling those of many ichneumonid ectoparasitoids such as the Tryphoninae and Cryptinae. However, many braconid endoparasitoids, s uch as the Microgastrinae, which emerge as final instar larvae from their host to complete feeding externally, also have similar mandibular teeth. In common with known ichneumonid endoparasitoids, lycorinine larvae: 1) lack a distinct labral sclerite, 2) have a disc-shaped antenna that lacks a central papillus, and 3) have an unsclerotized epistomal suture. The lack of a spiracular closing apparatus is apparently unique amongst the Ichneumonidae. Recent studies by Coronado-Rivera et al. (2004) and Shaw (2004) have partly resolved the issue. The eggs of three species have been examined and all possess a stalked anchor, very much like that found in the eggs of Tryphoninae; it is almost certainly used to attach the egg to the host cuticle. Coronado-Rivera et al. (2004) found that L. apicalis is a koinobiont that finishes its development as an ectoparasitoid but were unable to find evidence of early larval instars on the hosts exterior. Shaw observed that oviposition was into the hosts anus and concluded that it seems probable that the egg of Lycorina is positioned externally to the body of the host, albeit in a concealed site, and that Lycorinia [sic] is essentially an ectoparasitoid. (Shaw, 2004: 306). Shaw went on to postulate that it was unlikely Lycorina switched from endophagy to ectophagy, and that it is probably an external parasitoid throughout, owever long it may take for the larva to emerge from the hosts anus. (p. 307).
Diagnosis
Small to medium-sized ichneumonids, fore wing length 3.6 to 7.6 mm. Clypeus convex, separated from supraclypeal area by epistomal suture; apical margin medially trunctate to weakly concave, without denticles. Malar space with strongly impressed subocular groove. Mandible bidentate. Antenna slightly clavate apically; males without tyloids. Ventral posterior corner of propleuron without strongly produced lobe. Pronotum medially with strong but short longitudinal carina crossing transverse sulcus; epomia short, strong, with dorsal end produced as tooth-like projection. Mesoscutum with notauli absent. Mesopleuron with sternaulus absent, or short (extending 0.2-0.3 of way to middle coxa) and shallow. Posterior transverse carina of mesothoracic venter absent. Metanotum produced laterally into small hook that engages a protuberance on propodeum. Propodeum with carinae ranging from complete to absent; propodeal spiracles small and circular. Tarsal claws pectinate to apices. Fore tibia lacking a dorsal apical tooth. Fore wing with areolet absent (vein 3rs-m entirely absent); vein 2rs-m longer than abscissa of vein M between veins 2rs-m and 2m-cu; vein 2m-cu with a single short bulla. Hind wing with 1/Rs distinctly shorter than vein rs-m; veins 1-Rs, 1-M and 2-Cu nebulous to spectral (vein 2-cu sometimes basally incomplete). Metasomal insertion on propodeum between hind coxae. Metasoma depressed. Metasomal segment 1 with T1 broad anteriorly, posteriorly evenly broadened, with spiracles in front of midpoint and glymmae present; anterior sclerotized part of S1 short, not fused to T1. Thyridium of T2 present, short and wide (4-5x as long as wide), and adjacent to anterior tergal margin. T2-4 with narrow laterotergites separated by crease. T2-4 (and also less clearly T1) with impressed grooves delineating convex central triangular area. Female hypopygium large, in profile triangular, centrally membranous and with small median apical notch. Ovipositor projecting beyond apex of metasoma by more than length of hind tibia, slightly compressed; upper valve with distinct raised nodus, lower valve with four or more small teeth.
The Lycorininae present in the Nearctic
Lycorina
Holmgren, 1859 (HOD)
· Lycorina albomarginata (Cresson),
1874 (HOD)
· Lycorina
glaucomata (Cushman),
1920 (HOD)
·
Lycorina
scitula (Cresson), 1870 (HOD)
Identification and resources
The single genus Lycorina may easily be regognized in North America by the presence of a metapleural process and by the impressed triangular pattern on the metasomal tergites.
Key to Nearctic Species of Lycorina
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)