GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By Ian D. Gauld & David B. Wahl
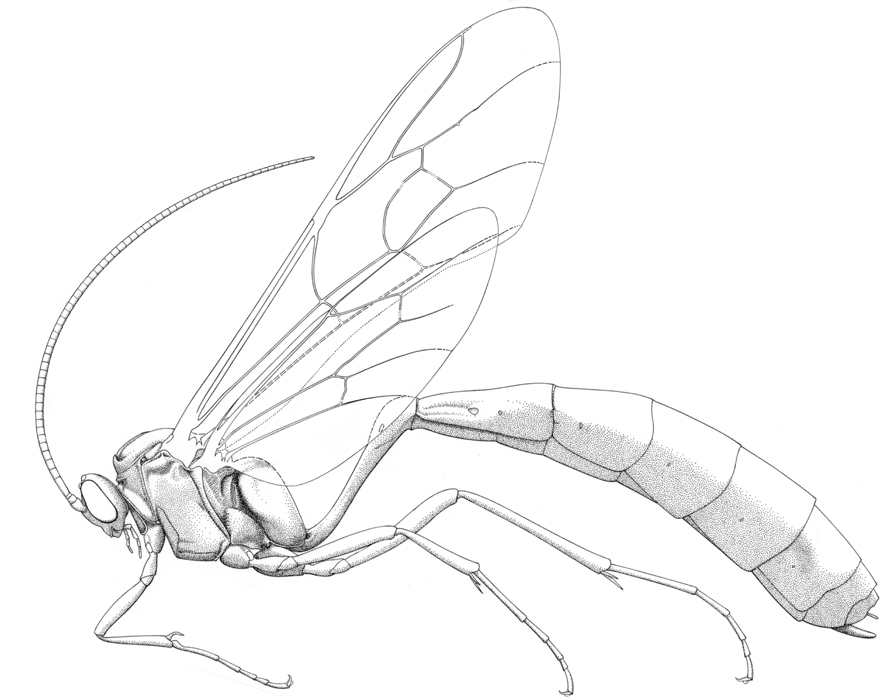
Thyreodon cyaneus |
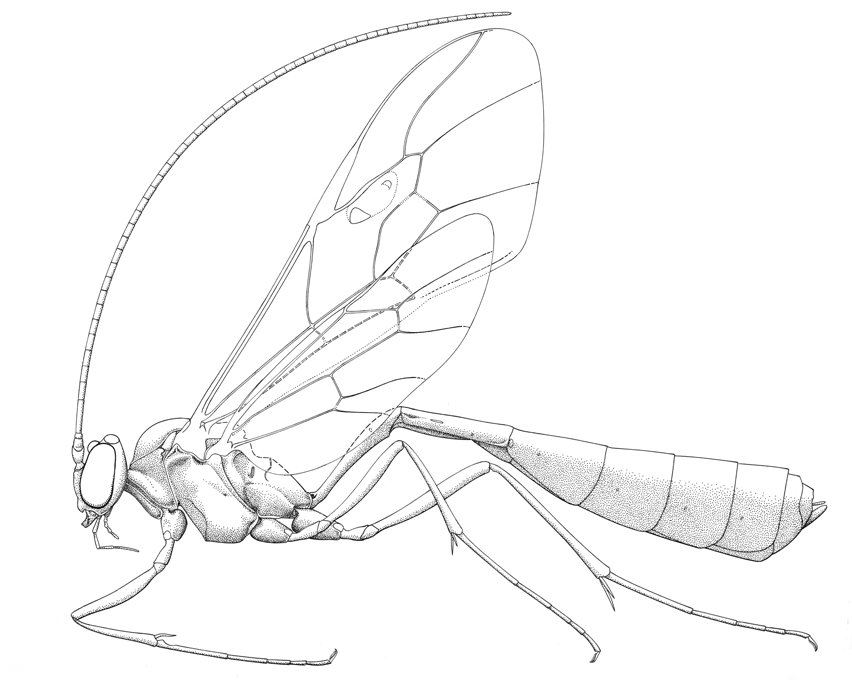
Enicospilus purgatus |
The Ophioninae includes the large orange-brown species that are commonly found at night flying around lights. All nocturnally active ichneumonids are pale, slender with long antennae and legs -- but not all are ophionines. Most commonly, species of the nocturnal tryphonine genus Netelia are mistaken for them, but these are readily distinguishable (see here). Ophionines can be found almost everywhere, and because they come to light they are easily collected in quite large numbers. Most museums and insect collections will have many specimens. Although the group is classified in 33 genera worldwide, the overwhelming majority of species encountered in the New World belong two genera: Ophion which predominates in temperate areas and on tropical mountain tops, and Enicospilus, a vast, primarily tropical genus. However, not all ophionines are nocturnal, and species of two day-flying genera, Thyreodon and Rhynchophion, occur in America. Both comprise mostly large, dark (blue, blue-black or brownish-red) species, many of which mimic aggressive aculeates such as the pompilid Pepsis. A few species of Thyreodon occur in North America, but the greatest diversity of the group is in tropical latitudes (Gauld & Janzen, 2004) where the group has apparently radiated to exploit the caterpillars of macroglossine Sphingidae and ceratocampine Saturniidae. Although the large dark species do not obviously closely resemble the nocturnal species, they are easily recognized as ophionines by the characteristic fore wing venation.
{kind=link}
Classification and diversity
There have been three major classifications proposed for the genera of Ophioninae (Cushman, 1947; Townes, 1971; Gauld 1985). The first author recognized three groups of genera, the Ophion, Thyreodon and Enicospilus genus-groups. Townes (1971) subsequently amalgamated Cushman's Thyreodon and Enicospilus generic groups and recognized two tribes, the Ophionini (= the Ophion genus-group) and the Enicospilini (= the Thyreodon + Enicospilus genus-groups). Gauld (1985) pointed out that whilst Townes's Enicospilini was monophyletic, his Ophionini was a paraphyletic assemblage. In order to create a more phylogenetic classification, Gauld arranged the 32 World genera of Ophioninae into five monophyletic genus-groups: the Ophion, Sicophion, Eremotylus, Thyreodon, and Enicospilus genus-groups. The first three genus-groups more or less correspond to the Ophionini sensu Townes, whilst the Thyreodon and Enicospilus genus-groups constitute that Enicospilini sensu Townes. The large Enicospilus genus-group was further divided by Gauld into five subgroups: the Orientospilus, Ophiogastrella, Stauropoctonus, Leptophion and Enicospilus genus-subgroups.
The Old World genera Hellwigia Gravenhorst and Skiapus Morley were treated as a tribe, Hellwigiini, in Campopleginae by Townes and later authors (Townes, 1970; Wahl, 1991) (but note that Horstmann (1969) raised the tribe to subfamilial status). Miah (1998) could not recover the tribe within the campoplegines, with his analysis indicating "its separate identity as a member of Ophioniformes group with affinities with Ophioninae and other subfamilies but not with Campopleginae." Quicke et al. (2005) recovered the two genera in or near the Ophioninae, never forming a monophyletic group. Hellwigia consistently nested deep within the Ophioninae in the combined morphological and molecular data sets, but Skiapus was the sister-group to Ophioninae; in the molecular data sets, Skiapus was erratic in its position. We accept Hellwigia as an ophionine but incertae sedis within the subfamily. Skiapus is problematical and cannot currently be assigned to a subfamily with any certainty. It should be treated as incertae sedis within the Ophioniformes.
Worldwide, there are 1025 described ophionine species (Yu & Horstmann, 1997), 48 of which are found in the Nearctic region (Carlson, 1979; Gauld, 1988a; Leblanc, 1989).
The general biology of Ophioninae
Ophionines are solitary koinobiont endoparasitoids of the larvae of other holometabolous insects. Virtually all rearings are from Lepidoptera, although a Nearctic species of Ophion is known to parasitize a coleopteran larva (Townes, 1971) and a European species has occasionally been recorded from a sawfly host (Thompson, 1957). The most usual hosts are moth larvae that feed exposed on vegetation, especially species of the families Noctuidae, Lasiocampidae, Lymantriidae, Saturniidae, Geometridae, Arctiidae, and Sphingidae. The larvae of Rhopalocera and Microlepidoptera are seldom attacked.
Adult ophionines occur in almost all habitats. Large numbers of species are crepuscular or strictly nocturnal (Rohlfs & Mack, 1985b). These insects are most frequently encountered at light, but sometimes they may be observed at dusk visiting flowers. Occasionally nocturnal species can be found roosting on the underside of leaves during the day. The diurnal ophionine species are more conspicuous and may often be encountered feeding from flowers or flying amongst shrub layer vegetation. Diurnally active koinobionts are probably exposed to a greater array of potential predators than are nocturnal species (Gauld, 1987), so it is not surprising to observe that many diurnal ophionines have protective devices. Some are aposematically coloured and resemble agressive aculeates. For example, Rhynchophion flammipennis is bluish-black with orange, black-tipped wings and in flight looks extremely like some species of Pepis (Pompilidae). Others, such as Thyreodon species, have elaborate flanges that guard the cervical membranes, and probably offer protection against asilid predators, whilst a few Thyreodon species have sharp processes on the mesoscutum which may serve to deter vertebrate predators.
Adult female ophionines are not known to feed on prospective hosts (Jervis & Kidd, 1986).
All the females I (IDG) have dissected (12 species) are apparently pro-ovigenic, that is they emerge from the pupa with a full complement of more or less mature eggs. Most individuals have between 7 and 30 mature eggs in each of their lateral oviducts. I have seen three individuals with one or no eggs present, suggesting further oocytes do not develop during active adult life. Rohlfs & Mack (1985a) observed that adult females of Ophion flavidus commenced oviposition within 24 hours of emerging from the pupa. Oviposition is into the host larva and is accomplished rapidly by the female straddling the host and thrusting her ovipositor forwards, into the host (Vickery, 1929). The ovipositor is withdrawn almost immediately. The ophionine egg is cylindrical, slightly curved, and pale coloured. No sign of paralysis has been observed in newly parasitized hosts. Many ophionines oviposit into relatively mature larvae in their third or fourth (Vickery, 1929), or fourth or fifth instar (Moutia & Courtois, 1952; Rohlfs & Mack, 1985a), though some species attack young larvae (Price, 1975). Generally a single egg is deposited free in the haemocoel. No species of ophionine are known to develop gregariously. It is not known if ophionines inject viral particles or physiology-manipulating venoms into their hosts. However, they may do so since species of the related Campopleginae are known to do both (Vinson & Stoltz, 1986), as are species in other subfamilies in the lineage to which the Ophioninae belong (Stoltz et al., 1981). The ichneumonid egg hatches to produce a caudate first instar larva (Clausen, 1940) that lives in the haemocoelic cavity of the host (Moutia & Courtois, 1952). In most cases it is not known what effect parasitization has on the host caterpillar, although parasitized hosts are sometimes, but not necessarily, smaller than healthy ones. One species of Ophion has been shown to depress host-larval food consumption (Rohlfs & Mack, 1983).
An ophionine may have three or more larval instars, though this facet of their biology requires closer investigation as it is notoriously difficult to assess the number of instars that endoparasitoids undergo (Rojas-Rousse & Benoit, 1977). Post first instar ophionine larvae are hymenopteriform in appearance (Clausen. 1940). The ophionine larva usually does not kill its host until after the host larva is fully grown, and often after it has spun a cocoon or constructed some other form of pupation retreat. The fully grown parasitoid larva eats out all the body tissue and fluids of the host, leaving only the cephalic capsule and cuticle (Vickery, 1929 and IDG pers. obs.). The final instar larvae of Ophioninae are typical endoparasites (Short, 1978): 1) the antenna is disc-shaped and lacks a central papillus, 2) the epistomal suture is unsclerotized, 3) the mandible is fairly small and without denticles on the blade, 4) the labral sclerite is absent, and 5) the spiracular closing apparatus is adjacent to the atrium. Ophionine larvae are unique for the combination of an elongate and ovoid labial sclerite, and numerous (20+) setae scattered over the labial region.
Generally the final instar larva destroys the host prior to its pupation, and the wasp larva then spins a thick cocoon within the host cocoon or in the host's pupation chamber in the soil (Rohlfs & Mack, 1985a). One Palaearctic species of Ophion is unusual in that it spins a cocoon within the host pupa (Brock, 1982), whilst an American species of Enicospilus kills its host as a fully grown larva and spins a cocoon within the unbroken host larval skin (Hancock, 1926). The cocoons are rather characteristic, being dark brown ovoids with a pale circumferential stripe. Like many other Hymenoptera (Gauld & Bolton, 1988), ophionines apparently have a brief pupal instar. A considerable amount of the time the ophionine spends within its cocoon may be passed as a final instar larva, and species which enter prolonged 'pupal' diapause probably do so as larvae. In north-western Europe one species of Ophion which emerges very early in spring passes the winter as a (?pharate) adult in the cocoon (Morley, 1915).
A great deal of work still needs to be undertaken on the host relationships of ophionines. It is very difficult to establish the real host range of any one species from the literature, as misidentifications of both parasitoids and hosts abound. Host lists of the type compiled by Thompson (1957) can be misleading as they simply collect all records, but fail to re-evaluate the evidence upon which each is based (see also Askew & Shaw, 1986). All too often one finds that several congeneric sympatric species of ophionine are listed as having identical or very similar host ranges. For example, in the U.S.A. the two fairly common large ophionines Enicospilus americanus and E. glabratus were frequently confused in early literature, and they are both recorded from a similar range of saturniid, arctiid and lymantriid hosts (Hooker, 1912). Closer study has suggested that the former species only attacks saturniid larvae, whilst the latter is restricted to the larvae of arctiids and lymantriids (Gauld, 1988a).
The evidence presently available indicates that there is a wide range of host specificity amongst species of Ophioninae. Many will attack a variety of hosts within a given family. The Central American species Thyreodon atriventris is believed to parasitize a range of sphingid larvae, but it has never been reared from similar-sized sympatric larvae belonging to other families (Janzen, pers. comm.). Other ophionines are more restricted in their host range, such as to hosts of a single subfamily; Thyreodon santarosae is only known to parasitize ceratocampine saturniids (Porter, 1986; Gauld, 1988b). Other species are believed to have an even more restricted host range and some apparently are monophagous. For example, Enicospilus lebophagus has only been reared from the saturniid Rothschildia lebeau despite intensive rearing of other sympatric possible hosts (Janzen, pers. comm.; Gauld 1988a).
Diagnosis
Medium-sized to large ichneumonids, fore wing length 6-29 mm. Labrum generally exposed. Clypeus usually weakly convex, separated from supraclypeal area by epistomal suture (suture rarely obsolescent); apex truncate, slightly concave, or convex, never with a median tooth or teeth. Mandible bidentate, sometimes twisted. Ocelli almost always large, posterior ones separated from eyes by less than their maximum diameter. Occipital carina usually complete, rarely completely absent. Antenna generally very long and slender, often with more than 55 flagellomeres; males without tyloids. Ventral posterior corner of propleuron without strongly produced lobe. Pronotum with epomia absent. Mesoscutum usually sparsely punctate; notauli weak to obsolescent. Sternaulus absent. Posterior transverse carina of mesothoracic venter complete or interrupted before middle coxa. Propodeal carinae varying from complete to having only anterior transverse carina present and the posterior part coarsely sculptured; very rarely without carinae. Legs slender. Fore tibia lacking a dorsal apical tooth. Tibial spurs very long, usually unequal. Tarsal claws long, pectinate to apices. Fore tibia lacking a dorsal apical tooth. Fore wing without areolet (2/Rs absent), vein 3rs-m distad vein 2m-cu; cell 3Cu with adventitious vein, originating at vein 2/1A, and paralleling wing margin; cell 1M+1R1 often with anterior glabrous area and sclerotized inclusions. Hind wing with vein 2/Rs straight to curved, longer than vein 1r-m; vein 2-Cu always present in New World species. Metasomal insertion on propodeum between hind coxae. Metasoma strongly laterally compressed. Metasomal segment 1 long and slender, T1 and S1 fused and without trace of tergosternal suture; spiracles near apex and glymmae absent. Ovipositor short and straight, about as long as apical metasomal depth (longer only in a few Old World species); dorsal subapical notch usually present. Body color usually brownish-red to whitish-yellow, sometimes with slightly darker maculation on mesosoma.
{kind=link}
{kind=link}
The genera of Ophioninae present in the Nearctic Region
- Enicospilus Stephens, 1835. Worldwide; 23 described Nearctic spp. (HOD)
- Eremotylus Förster, 1869. Holarctic; five described Nearctic spp. (HOD
- Ophion Fabricius, 1798. worldwide except Ethiopian and Australian; eleven described Nearctic spp. (HOD)
- Rhynchophion Enderlein, 1912. Nearctic, Neotropical; one described Nearctic sp. (flammipennis (Ashmead)). (HOD)
- Simophion Cushman, 1947. Holarctic; one described Nearctic sp. (excarinatus Cushman). (HOD)
- Thyreodon Brullé, 1846 (incl. Athyreodon Ashmead, 1900). Nearctic, Neotropical; six described Nearctic spp. (HOD)
- Trophophion Cushman, 1947. Nearctic; one described Nearctic sp. (tenuiceps Cushman). (HOD)
{kind=link}
Identification and resources
The majority of ophionines are easily recognized as they are moderately large, brownish-red to whitish-yellow wasps with large ocelli, long legs and long slender antennae. However, they may be confused with other nocturnal ichneumonoids, but are easily separable by several obvious features, especially of wing venation. A few species of ophionines belonging to Thyreodon and Rhynchophion are diurnal and blackish in color, but these may be recognized by their characteristic venation as described in the diagnosis above.
Key to New World
Genera of Ophioninae
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)
Currently there are no modern keys to the majority of Nearctic species, except for a species-group of Enicospilus (Gauld,1988a) and a species-group of Eremotylus (LeBlanc, 1989).