GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By Ian D. Gauld & David B. Wahl
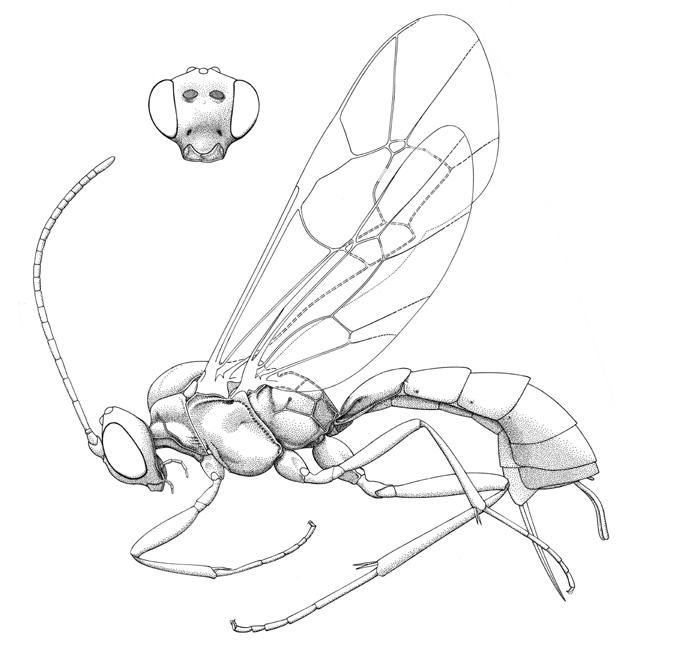
Stilbops vetula |
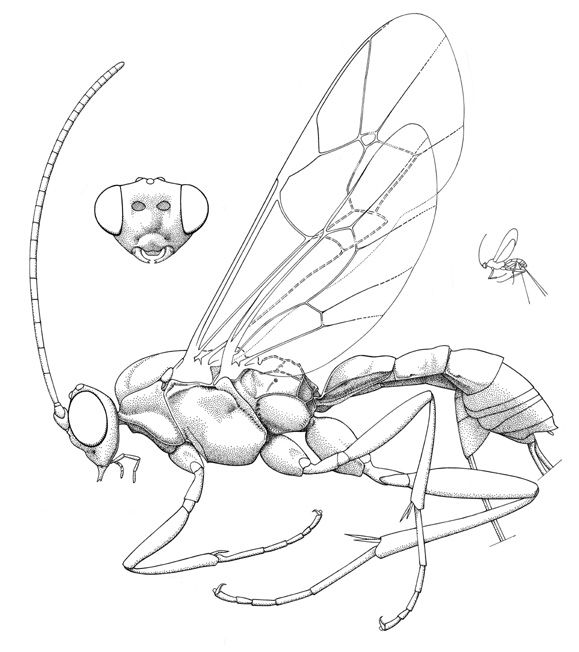
Panteles schutzeana |
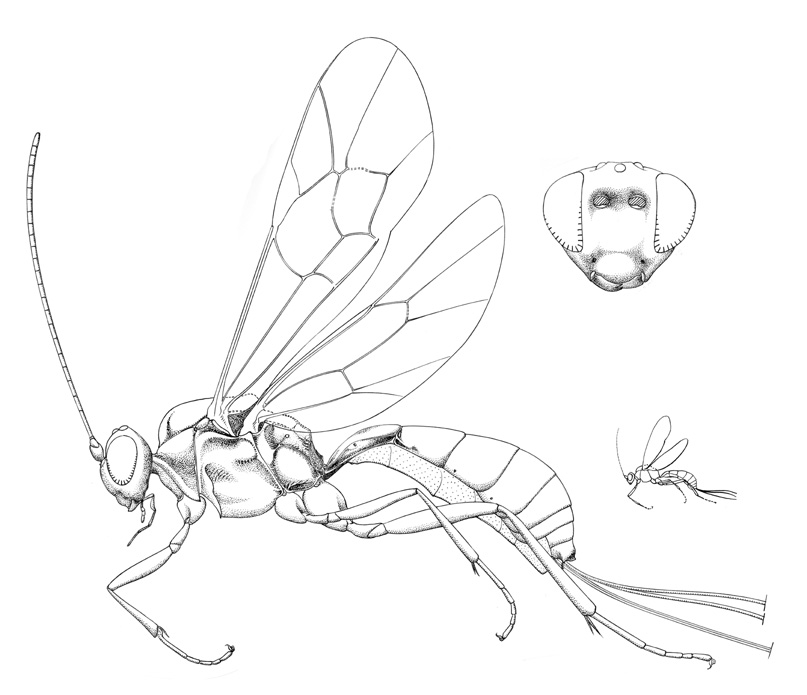
Notostilbops fulvipes |
Except for the European species Stilbops vetula (Gravenhorst), stilbopines are rather uncommon ichneumonids found only in temperate habitats. Unlike many other primarily temperate subfamilies, the Stilbopinae is amphitropical with two small genera, Stilbops Foerster and Panteles Foerster, in the Holarctic and one, Notostilbops Townes, in the temperate south of Chile (Townes, 1970). Stilbopines superficially resemble atrophine Banchinae, but all species have a complete complement of propodeal carinae which define an areola. Stilbopines are solitary koinobiont endoparasitoids of Incurvariidae (Lepidoptera), although nothing is yet known of the biology of Notostilbops.
Classification and diversity
The Stilbopinae is among the smaller subfamilies of Ichneumonidae, comprising three genera (Yu & Horstmann, 1997; Kasparyan, 1999): Stilbops with 21 described species in the Holarctic, the northern Neotropic region and the northern periphery of the Oriental region; Panteles with one species in Europe and a second species in the Kuril Islands; and Notostilbops with a single described species in southern Chile.
The classification of Stilbops has been controversial. Townes (Townes & Townes, 1951) placed the genus in a separate tribe in the Tryphoninae. He subsequently (Townes, 1970) united Stilbops with Panteles and Notostilbops, placing the expanded Stilbopini as a tribe of Banchinae. Short (1978) elevated Stilbops to the rank of a separate subfamily (Stilbopinae) on the basis of the morphology of the mature larva; he did not address the status of Notostilbops or Panteles. Townes later (Townes & Townes, 1978) included Notostilbops in Stilbopinae but transferred Panteles to the Atrophini. However, Wahl (1988) placed Panteles back in the Stilbopinae and stated that the Stilbopinae appear to be a monophyletic group of endoparasitoids unrelated to the Banchinae (or Ctenopelmatinae or Ophioninae s.l.). Wahls interpretation has been followed by subsequent authors (Gauld, 1991; Yu & Horstmann, 1997; Quicke, 2005).
The general biology of Stilbopinae
Stilbops and Panteles are known to be parasitoids of the Incurvariidae, a basal, non-ditrysian family of Lepidoptera (Grimaldi & Engel, 2005). Incurvariids are rather unusual hosts for Lepidoptera-parasitizing ichneumonids, which almost exclusively utilize Ditrysia.
Various Palaearctic species of Stilbops are known to be egg-larval parasitoids of Adelinae (Adela and Nemophora) with the female parasitoid ovipositing into the moths eggs and the final instar larval ichneumonid killing its host in the moths cocoon (Hinz, 1981). One species with an unusually long ovipositor, S. ruficornis (Gravenhorst), has been observed to probe within the flower heads of Knautia arvensis (L.) Coulter (Dipsaceae) for the concealed eggs of Nemophora metallica (Poda) (Fitton, 1984). Stilbops vetula uses its much shorter ovipositor to parasitize the eggs of Adela reaumurella (L.), which are placed in pockets in the epidermis of various plants. After a short feeding period, the moth larvae drop to the ground, construct cases from leaf fragments and feed dispersed in the leaf litter. Panteles schuetzeanus (Roman) is an endoparasitoid of the gall-forming incurvariid Lampronia fuscatella (Tengström) (Quicke, 2005), and this author suggests that the ichneumonid probably gains ovipositional access to its host by boring through the wall of the woody gall. The parasitoid egg is rather large (0.45 mm long), with a dark cuticle and a curved hook-like appendage at one end; it is placed in the host larva (generally in the caudal region), with the curved hook embedded in the host tissue. Although superparasitism was common, only one egg hatches per host, with the development of other eggs apparently being chemically suppressed. After killing its host, the Panteles larva spins a thin silken cocoon within the capped Lampronia gall (Quicke, 2005).
Although it is easy to emphasize the striking differences between of Panteles and Stilbops, and to question whether or not they are correctly associated (as did Townes & Townes, 1978 and Quicke, 2005), these differences can largely be explained by adaptation to differences in the biology of the host incurvariids (Heath & Pelham-Clinton, 1976). Since Stilbops oviposits into the host egg, the ovipositor is finely pointed and the egg is small and lightly sclerotized. Panteles, however, attacks a gall-forming host and has an ovipositor with dorsal subapical teeth more suitable to drilling through woody tissue; the egg is larger, darkly pigmented, and placed in moderately well-grown larvae. It is noteworthy that similar morphological differences occur in the campoplegine genus Venturia, between species in the ovivena complex (Zwart, 1973), which are egg/larval parasitoids, and those species in the nigriscapus group (Wahl, 1984) which are larval parasitoids.
Diagnosis
Small ichneumonids, fore wing length 3.8 to 5.2 mm. Labrum hidden or exposed (Stilbops), apical margin convex. Clypeus varying from convex to apically flattened, weakly separated from supraclypeal area by epistomal suture; apical margin usually convex, without denticles. Malar space without subocular groove. Mandible bidentate. Antenna with flagellum filiform; males without tyloids. Ventral posterior corner of propleuron without strongly produced lobe. Pronotum with epomia present dorsally. Notauli weakly to strongly impressed, extending about 0.3 length of mesoscutum. Epicnemial carina present. Sternaulus present, extending 0.2-0.4 of way to middle coxa. Posterior transverse carina of mesothoracic venter present only as central vestige. Submetapleural carina of metapleuron widened anteriorly into flange. Propodeum with complete complement of carinae; propodeal spiracles small and circular. Tarsal claws ranging from simple to basally pectinate to completely pectinate. Fore tibia lacking dorsal apical tooth. Fore wing with areolet present and obliquely quadrate, or absent (vein 3rs-m entirely absent); vein 1cu-a distad vein M by 0.5 or more times its own length; vein 2m-cu with one (Panteles) or two bullae. Hind wing with 1/Rs equal to or greater than vein rs-m; veins 1/Rs, 1/M and 2/Cu nebulous to spectral. Metasomal insertion on propodeum between hind coxae. Metasoma basally dorsoventrally depressed, apical 0.3-0.5 laterally compressed. Metasomal segment 1 with T1 evenly broadened posteriorly, with spiracles in front of midpoint and glymmae present; anterior sclerotized part of S1 short, not fused to T1. Thyridium of T2 present, transversely ovoid (2-3x as wide as long), and adjacent to anterior tergal margin. T2-3 with laterotergites separated by crease, laterotergite of T4 separated or pendant. Female hypopygium large, in profile triangular, apex with small median apical notch (Panteles) or median membranous area (Notostilbops and Stilbops). Ovipositor either projecting beyond apex of metasoma by more than length of hind tibia (Notostilbops and Stilbops) or equal to or less than length of hind tibia (Stilbops); apex variable: 1) apex not tapered to sharp point, dorsal valve with subapical notch, apex of ventral valve without teeth (Notostilbops), 2) apex not tapered to sharp point, dorsal valve with three subapical teeth dorsal and no subapical notch, apex of ventral valve without teeth (Panteles), 3) apex gradually tapered to sharp point, dorsal valve without subapical notch, apex of ventral valve without teeth (Stilbops).
The Stilbopinae present in North and Central America
A single species, Stilbops latibasis Townes, is present in North America. A second species, S. mexicana Townes, is present in Mexico (Townes, 1989). Nothing is known of the biology of either species.
Stilbops Förster, 1869 (HOD){kind=link}
{kind=link}
Identification and resources
Key to species of New World Stilbops
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)
Female Stilbops are easily recognized by the combination of the:
- sharply tapered ovipositor and large hypopygium,
- strong dense punctation on T2-4,
- fore wing with vein 1cu-a distad vein M by 0.5 or more than its own length, and
- head morphology (large anterior tentorial pits; clypeus with basal 0.4 convex, apical 0.6 flattened or weakly concave, and apical margin strongly convex).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Males are more difficult and beginners are likely to confuse them with male pimplines (particularly Ephialtini). The combination of the aforementioned head characters, metasomal punctation, and venational character of the fore wing will give the best chance of recognition.