GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By David B. Wahl & Ian D. Gauld
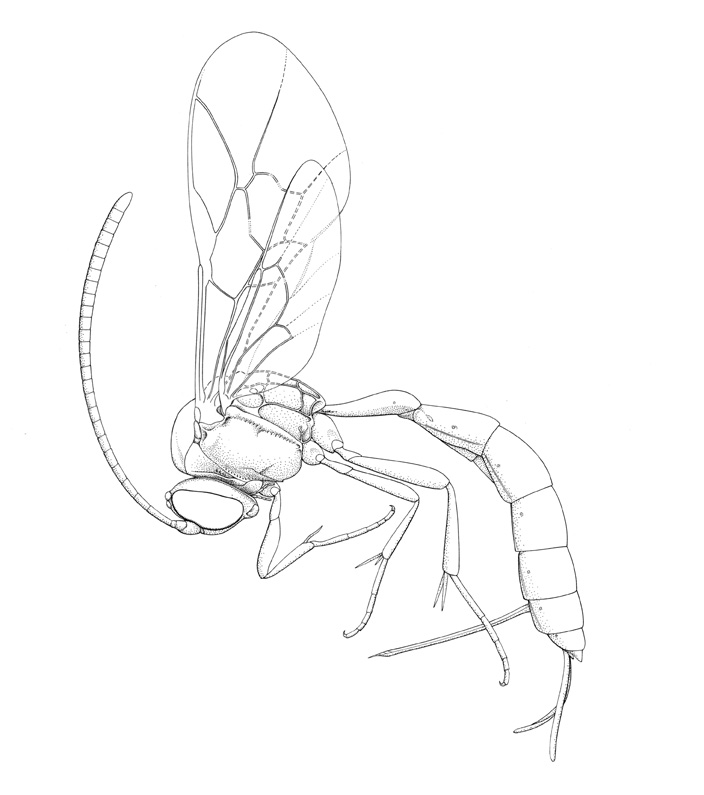
Brachycyrtus ornatus |
The subfamily Brachycyrtinae is a small group of uncommon ichneumonids placed in a single genus, Brachycyrtus. They are amongst the most easily recognized of all ichneumonids on account of their characteristically modified fore wing venation (vein 1cu-a is far distad the base of vein Rs&M), elongate first metasomal segment inserted above the hind coxae, and color (usually yellow with extensive black markings). In the New World, the greatest species richness occurs in South America, but two species are present in North America.
Classification and diversity
The modem concept of the Brachycyrtinae originated with Townes (1969), who placed four genera (Adelphion, Brachycyrtus, Pedunculus and Poecilocrytpus) together for the first time as a tribe in the Labeninae. Gauld (1983) removed Poecilocrytpus to a separate new tribe, and subsequently (Gauld, 1984) added a new Australian genus, Monganella, to the Brachycyrtini. A decade later Wahl (1993) elevated the group to the status of a distinct subfamily, Brachycyrtinae. This new status has been widely accepted in subsequent work (e.g., by Hanson & Gauld, 1995; Gauld & Wahl, 1999) even though doubts existed about the integrity of the group. Although there are some similarities between the included genera, for example in the form of metasomal segment 1 in which the tergite and sternite are fused into a seamless cylinder, the monophyly of the group had not been unequivocally demonstrated. Without analysis or any clear explanation, Porter (1998), in his treatment of the Chilean fauna, placed Pedunculus in a separate new subfamily, the Pedunculinae. Furthermore, Porter created some confusion as he failed to state whether the other two Australian genera belonged in his new group, or remained in the Brachycyrtinae. Gauld & Ward (2000) re-examined all four genera and failed to find any convincing synapomorphies uniting them, thus concurring with Porter's action of placing Brachycyrtus and Pedunculus in separate subfamilies. Additionally, they concluded that Adelphion and Monganella are best placed with Pedunculus in the Pedunculinae, and that both Brachycyrtinae and Pedunculinae are monophyletic. Gauld & Ward listed the more important differences between these two subfamilies (Gauld & Ward, 2000: 15). The Brachycyrtinae is thus restricted to contain only the single genus Brachycyrtus.
Brachycyrtus is a cosmopolitan genus comprising 21 described species, found mainly in the tropics, including remote oceanic islands (Yu & Horstmann, 1997; Gauld & Ward, 2000). Two species are present in the Nearctic region.
The general biology of Brachycyrtinae
Brachycyrtus species appear to be solitary ectoparasitoids of the cocooned pupae and prepupae of Chrysopidae (Neuroptera). All reared specimens examined by Gauld & Ward (2000) are from such hosts. Although there are occasional records of other hosts, these need verification before they can be accepted. It is, however, worth pointing out that in the New World all of the species known to attack Chrysopidae belong to a single morphologically derived species-group, the ornatus species-group (Gauld & Ward, 2000). None of the structurally more diverse species have been reared. In the United States, Brachycyrtus pretiosus has been reared from the cocooned prepupae of the chrysopid Nodita pavida (Hagen) (Cushman, 1936), and there are records of other species from green lacewings in other areas.
The final instar larvae of Brachycyrtus possess typical ectoparasitoid features (Short, 1978; Wahl, 1993): 1) the mandibular blade is denticulate on the dorsal margin, 2) the labral sclerite is present, and 3) the spiracular closing apparatus separated from the atrium by a section of trachea. The antenna has not been observed in the various slide-mounted preparations; since no ectoparasitoid is known to have a disc-shaped antenna, determination of its condition would be most interesting.
Diagnosis
Small to medium-sized ichneumonids, fore wing length 3.1-9.0 mm. Labrum exposed beyond apical margin of clypeus. Clypeus small and rather flat, usually not clearly separated from face by epistomal suture; apex thin, varying from truncate to slightly convex, without any trace of median or lateromedian apical teeth. Mandible bidentate, generally stout and rather short, frequently with apex of dorsal tooth impressed, so mandible appears to be tridentate (as in the Diplazontinae). Supra-antennal area flat to weakly concave, without median vertical carina. Occipital carina present, meeting hypostomal carina either above or at mandibular base. Antenna with flagellum slender to distinctly clavate apically; males without tyloids. Mesosoma generally rather short and deep. Ventral posterior corner of propleuron with strongly produced lobe. Pronotum with prominent vertical epomia, apex ranging from below midheight to almost touching dorsal margin. Mesoscutum with notauli usually absent, rarely present; when present then about as long as width of lateral ocellus, sometimes with a small crest anteriorly. Sternaulus short (extending 0.2-0.4 of way to middle coxa) or absent. Posterior transverse carina of mesothoracic venter usually complete, rarely broadly incomplete. Propodeum short and steeply declivous posteriorly, generally with complete set of carinae (median section of posterior transverse carina usually absent); propodeal spiracles small and circular. Tarsal claws simple. Fore tibia lacking a dorsal apical tooth. Apex of hind tibia posteriorly with fringe of stout setae. Middle and hind tibial spurs inserted in an area confluent with the basitarsus.. Fore wing with a transversely pentagonal areolet that lacks vein 3rs-m; vein 1cu-a distad vein Rs&M by = 0.8x its own length; vein 2m-cu with two bullae. Hind wing with vein 1/Rs conspicuously shorter than vein rs-m; vein 2-Cu spectral. Metasomal insertion on a short neck, distinctly above insertion of hind coxae. Metasoma moderately stout, slightly dorso-ventrally compressed. Metasomal segment 1 with T1 and S1 usually entirely fused and without trace of a suture, moderately slender, postpetiole only slightly wider than petiole; spiracles near apex and glymma absent. Thyridium of T2 present, elongate and narrow, anterior margin touching tergal base. T2-3/4 with broad laterotergites separated by crease. Ovipositor projecting beyond apex of metasoma by 1-2x length of hind tibia; upper valve without a dorsal subapical notch, lower valve without teeth. Male usually with aedeagus moderately stout, subcylindrical, apically rounded.
The Brachycyrtinae present in the Nearctic
Brachycyrtus Kriechbaumer, 1880 (HOD)
· Brachycyrtus ornatus Kriechbaumer, 1880 (HOD)
· Brachycyrtus pretiosus Cushman, 1936
(HOD)
Identification and resources
Species of Brachycyrtus are easily recognized by the characteristic venation of the fore wing as described above. Although superficially resembling a small cryptine, no cryptine has vein 1cu-a distad vein Rs&M by 0.8x its own length as does Brachycyrtus.
Key to species of Nearctic Brachycyrtus
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)