GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By Ian D. Gauld & David B. Wahl
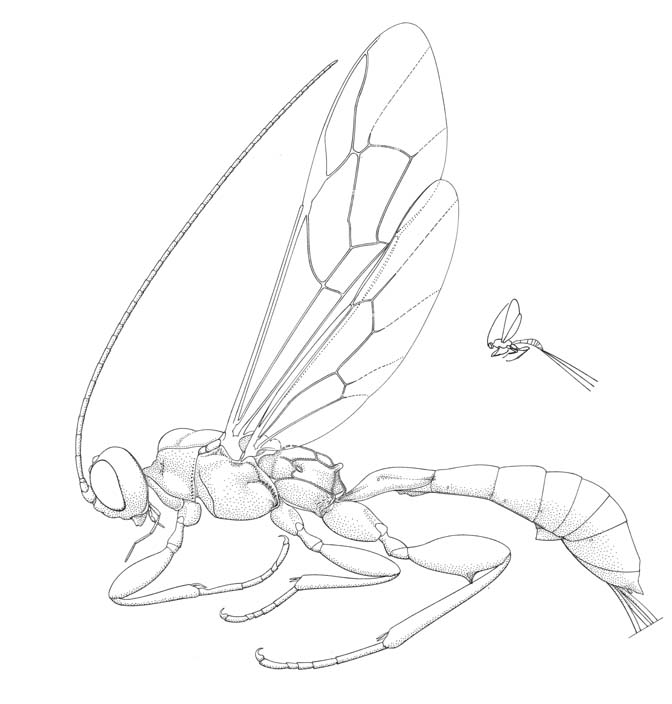
Odontocolon ochropus |
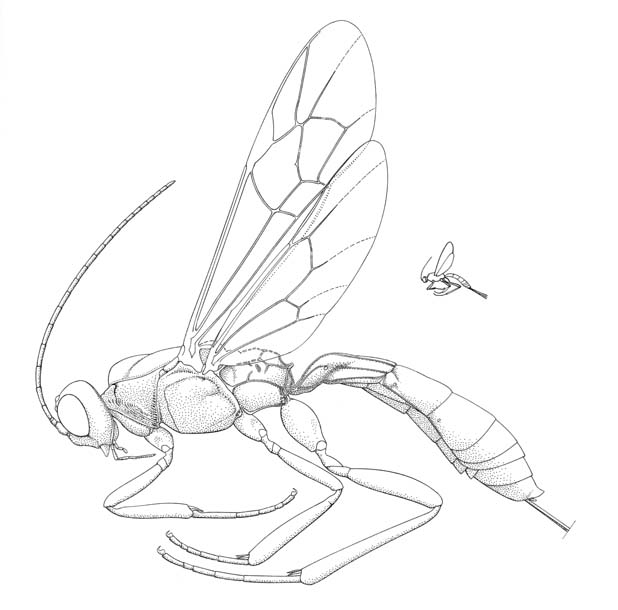
Xorides irrigator |
The Xoridinae comprises a group of rather rare, quite elongate ichneumonids that are most commonly encountered in mature growth forests where they search for their beetle hosts in moribund timber. Like other parasitoids of wood-boring insects female, xoridines possess a long conspicuous ovipositor. Although never very common, four genera are found throughout North America; in the tropical regions to the south only one genus, Xorides, is encountered.
Classification and diversity
The Xoridinae is one of the smaller subfamilies of Ichneumonidae, with 204 described species for the World (Yu & Horstmann, 1997), classified into four genera (Townes, 1969). Three of these, Ischnoceros, Odontocolon and Aplomerus, are confined to the North Temperate region (or to the interface of the Palaearctic with the Oriental region), whilst the fourth, Xorides is cosmopolitan. Wahl (1997) analyzed the cladistics of the genera and subgenera; the subgenera of Xorides were reduced to species-groups.
The general biology of the Xoridinae
Xoridines are idiobiont ectoparasitoids of wood-boring Coleoptera and Symphyta. Larvae usually serve as hosts, but pupae and even adults within their cocoons or pupation chambers may be parasitized. Species of the families Cerambycidae and Siricidae are the commonest recorded hosts (e.g. Chrystal & Skinner, 1931; Sharifi & Javadi, 1971), though there are isolated records from other coleopterous families, and some species are at least facultatively hyperparasitic (Austin et al., 1994). Most host data pertains to European and North American species and there are virtually no host records from the tropics.
There are a number of studies of the biology of xoridines in Europe and North America. That of Chrystal and Skinner (1931), on the biology of Xorides species attacking the cerambycid, Tetropium gabrieli, in conifers in Europe serves as a typical example. These authors observed that the males tended to emerge first and search around the bark of suitable trees waiting for the females to emerge. The male approached the female with his antennae vibrating rapidly and mounts her from behind, curling his metasoma under hers. Pairing may last from 15 to 40 minutes and during this time he continued to stroke the female with his antennae. The antennae of many male xoridines are rather highly modified - often with elongate fringes of hairs, and presumably have an important role in courtship.
The female xoridine oviposited through the bark into the host gallery a process observed to take between 15 and 40 minutes - rarely sometimes much longer. Chrystal and Skinner comment that the degree of success and accuracy of oviposition seemed to be far greater than that of Rhyssa. A single egg was deposited on or near the cerambycid larva, and there is some evidence for host specificity as in trials the ichneumonid had little interest in other species of cerambycid. Although it was never observed, Chrystal and Skinner considered it was highly likely that the host larva was stung and paralysed prior to oviposition. Larvae that had been oviposited on were found to be immotile, but alive, and even if the xoridine egg failed to hatch the larva remained unmoving for several weeks before eventually dying.
The elongate, fusiform egg hatched about eight days after oviposition, and the first instar larva immediately began to attack the host. The first instar larva was observed to be more or less conical, with a well-developed cephalic capsule and prominent mandibles. It fed by burying these mandibles in the host's integument and sucking the body fluids. Growth was rapid and after six or seven days the larva moulted. The second instar larva resembled the first and fed for a further four or five days before moulting again. At about this time feeding resulted in the death of the host. The third instar larva was stouter than the preceding two with a less strongly developed head. This continued to feed on the dead host for a further seven or eight days before moulting. The fourth and final instar larva consumed the entire host except for its head capsule and chitinous exoskeleton in five or six days. The larva then remained motionless for a few days before spinning a capacious cocoon. The larva was observed to overwinter in this cocoon and pupated in the spring of the following year. The duration of the pupal stage was short, with the adult emerging after nine or ten days.
The final instar larvae of Xoridinae are typical ectoparasitoids (Short, 1978): 1) the antenna has a central papillus, 2) the epistomal suture ranges from partially sclerotized to unsclerotized, 3) the mandibular blade is denticulate on the dorsal margin, 4) the labral sclerite is present, and 5) the spiracular closing apparatus separated from the atrium by a section of trachea.
Diagnosis
Small to large ichneumonids, fore wing length 2.8 to 25.0 mm. Clypeus quite small, separated from supraclypeal area by epistomal suture, usually with strong transverse ridge and with flattened apical area; apex truncate or convex. Malar space with a weakly to strongly impressed subocular groove. Mandible short, apex with one or two teeth. Supra-antennal area sometimes with crest between antennal sockets. Antenna with flagellum filiform; males without tyloids. Ventral posterior corner of propleuron without strongly produced lobe. Prontoum with epomia varying from absent, to long and strongly developed, projecting dorsally as a tooth. Notauli strongly impressed, generally more or less meeting centrally and extending posteriorly towards scuto-scutellar groove. Sternaulus absent or short. Posterior transverse carina of mesothoracic venter usually present as a central vestige or absent (complete only in some Old World species of Xorides). Propodeum long, generally with complete set of carinae, usually with apophyses well-developed. Tarsal claws simple. Fore tibia with sub-basal constriction; dorsal surface with denticles; dorsal apical tooth absent. Hind femur swollen. Fore wing without areolet, vein 3rs-m entirely absent, vein 2/Rs shorter than abscissa of vein M between veins 2/Rs and 2m-cu; vein 2m-cu with two short widely interspaced bullae. Hind wing with vein 1/Rs distinctly shorter than vein rs-m; veins 1-Rs, 1-M and 2-Cu tubular to nebulous. Metasomal insertion on propodeum between hind coxae. Metasoma cylindrical or dorsoventrally depressed. Metasomal segment 1 with T1 narrow anteriorly, posteriorly evenly broadened, with spiracles at or in front of midpoint and glymmae absent; anterior sclerotized part of S1 short, usually fused to T1 (free in Aplomerus). T2-4 smooth or with weakly impressed grooves. Hypopygium of female short, transverse. Ovipositor projecting beyond apex of metasoma by at least length of hind tibia and frequently longer; upper valve without dorsal subapical notch, apex of lower valve with conspicuous teeth.
The genera of Xoridinae present in the Nearctic Region
- Aplomerus Provancher, 1886; Nearctic, Neotropical, Palaearctic; five described Nearctic species. (HOD)
- Ischnocerus Gravenhorst, 1829; Holarctic, Palaearctic/Oriental interface; one described Nearctic species (clivulus Townes). [One species is purported to occur in southern South America (lehmannnitschei Schrottky, 1902) but the unique specimen is now lost and the record is currently unverifiable.] (HOD)
- Odontocolon Cushman, 1942; Holarctic, Palaearctic/Oriental interface; 23 described Nearctic species. (HOD)
- Xorides Latreille, 1809; Worldwide; 21 described Nearctic species. (HOD)
{kind=link}
Identification and resources
All four genera are represented in the Nearctic fauna and a key to these is accessible below. Keys to species are in Townes & Townes (1960).
Key to genera of World Xoridinae
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)