GIN Home
Ichneumonid Morphology
Subfamily Key
Lists of World Genera
Acaenitinae
Brachycyrtinae
Collyriinae
Lycorininae
Ophioninae
Poemeniinae
Rhyssinae
Stilbopinae
Xoridinae

By Ian D. Gauld & David B. Wahl
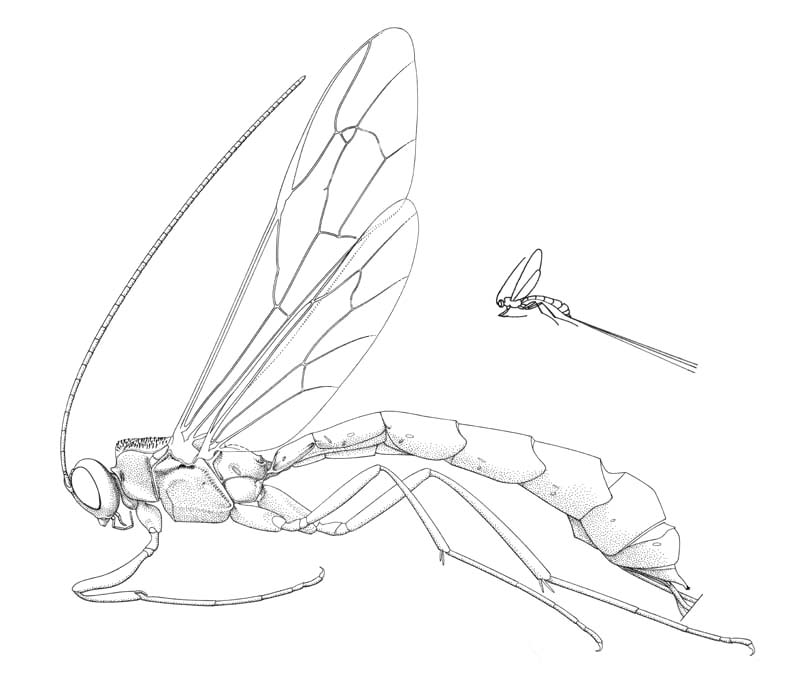
Megarhyssa nortoni |
The subfamily Rhyssinae includes some of the most spectacular of all Ichneumonidae. These large conspicuous wasps, the females of which have very long ovipositors and may exceed 200 mm in length, are quite commonly encountered in the northeastern states and western forests, but individuals seem progressively rarer in the southern United States. Rhyssines are parasitoids of the wood-boring larvae of Siricoidea, and female rhyssines aggregate around siricoid-infested trees where they may spend long periods boring into the wood to gain ovipositional access to their hosts. Occasionally aggregations of males may be found around trees from which females are in the process of emerging (Eggleton, 1990).
Classification and diversity
The Rhyssinae is a small cosmopolitan subfamily that is perhaps most species-rich in the lowland tropical forests of southeast Asia (Kamath & Gupta, 1972). Worldwide, it includes 234 described species in eight genera (Yu & Horstmann, 1997). Until quite recently (e.g. Townes, 1969) the rhyssines were treated as a tribe within the Pimplinae, but this was based upon shared primitive features (Eggleton, 1989). Their many unique derived features suggest rhyssines are better treated as a distinct subfamily, and the Rhyssinae and Pimplinae are currently treated as sister groups within the Pimpliformes (Gauld, 1991; Wahl & Gauld, 1998).
In the New World, the highest rhyssine generic diversity occurs in the northeastern hardwood forests, whilst further south in tropical countries there are only two genera Rhyssa and Epirhyssa. The former genus is represented in the mountains of Honduras by a single species (Porter, 2001), but the latter genus is species-rich throughout tropical America (Porter, 1978; Gauld, 1991). Recent work shows a very high species-richness of Epirhyssa in Amazonian lowland forest, and fails to uphold Porters (1978) hypothesis that the greatest species-richness of rhyssines occurs on the middle slopes of the Andes (Sääksjärvi et al., 2004). Recently, a positive correlation has been found in Amazonia between rhyssine species-richness, and plant species-richness (Sääksjärvi et al., 2006).
The general biology of Rhyssinae
Rhyssines are idiobiont ectoparasitoids of the immature stages of endopterygote insects boring in wood. In the north temperate region, their usual hosts are the larvae of siricoid woodwasps (Siricidae and Xiphydriidae), but we suspect other wood-boring hosts are used in the lowland tropics where siricids are almost absent (totally so in tropical America), and xiphydriids seem to be exceedingly rare. In addition to being primary parasitoids of woodwasps, rhyssines may also develop as facultative hyperparasitoids using other woodwasp parasitoids as hosts (Hanson, 1939). Like many other idiobiont ectoparasites of wood-boring insects, rhyssine larvae are probably able to develop on virtually any endopterygote and some have even been cultured in the laboratory on entirely unnatural surrogate hosts (Spradbery, 1968).
Female rhyssines are highly specialized for drilling in timber: they have an extensive membranous area between tergites VI and VII which allows them to rotate the tip of the metasoma, a long rigid and more or less straight ovipositor, and several sternal tubercles that apparently act as an ovipositor guide (Gardiner, 1966). Females of some temperate species locate their hosts by responding to the presence of the siricid hosts fungal symbiont (Spradbery, 1970; Spradbery & Kirk, 1978) and this may result in a fairly large number of rhyssines being present on a single tree. At least some species are capable of drilling through several centimeters of hard wood to reach their host, an act that may take a considerable time (Morgan & Stewart, 1966). When their ovipositor is deep in the wood these insects are very vulnerable to avian predation as ovipositor withdrawal also takes some time. Having located a host, the rhyssine female apparently stings and paralyses it before laying a fully-yolked, large egg on it (Chrystal, 1937). The eggs of rhyssines are extremely elongate (Iwata, 1958) to allow them to pass down the lumen of the ovipositor. The parasitoids egg ecloses rapidly (Nuttall, 1980) and the larva commences to feed on the host via an integumentary lesion. When fully-fed, which is generally when it has consumed all but the hardest parts of the host, the rhyssine larva spins a loose brownish cocoon. It may then pupate, or delay development by diapausing as a larvae for up to a year (Nuttall, 1980).
Like most other Hymenoptera, the males of Rhyssinae often emerge from their pupation site a day or two before the females (Nuttall, 1980). Males of some temperate species, such as Megarhyssa nortoni have extremely elongate metasomas, whilst males of many tropical species have short, stout metasomas. Eggleton (1989) suggested these different male morphologies are the consequence of different selective pressures resulting from alternative types of mating behaviour scramble competition polygyny versus female defense polygyny (Thornhill & Alcock, 1983). Males are known to aggregate around logs from which females are emerging, and the stout males establish and defend territories where a female is likely to emerge (Eggleton, 1989). In some temperate species, scramble competition for females occurs, and in such situations any adaptation that allows a particular male to copulate before his fellows is likely to be selected for. Some males of these species have remarkably elongate metasomas that are inserted into the females emergence tunnel enabling the male to copulate with the female prior to her emergence from the log (Nuttall, 1973).
Diagnosis
Medium-sized to large ichneumonids, fore wing length 6.0 to 30.0 mm. Clypeus small, quadrate, and flattened; separated from supraclypeal area by epistomal suture; apex truncate or weakly concave, with median tubercle and/or lateral tubercles. Malar space without subocular groove. Mandible short and stout, apex bidentate, dorsal tooth often broadened and chisel-shaped. Occipital carina medio-dorsally incomplete. Antenna with flagellum filiform; males without tyloids. Gena without denticulation. Foramen magnum not laterally expanded. Ventral posterior corner of propleuron without strongly produced lobe. Pronotum with epomia present, not ventrally continued on anterior pronotal margin. Mesoscutum with prominent transverse rugae. Notauli strongly impressed, generally more or less meeting centrally and extending posteriorly towards scuto-scutellar groove. Sternaulus absent. Epicnemial carina usually present. Posterior transverse carina of mesothoracic venter absent. Propodeal carinae absent; propodeal spiracle = 2.0x as long as wide. Tarsal claws simple. Fore tibia with dorsal apical tooth absent. Hind coxa elongate, length = 2.3x width. Fore wing with areolet present (petiolate triangular) or absent; vein 2m-cu with two bullae. Hind wing with vein 1/Rs = vein rs-m; veins 1-Rs, 1-M and 2-Cu tubular; vein 2/Cu very close to, or appearing to originate from, vein M. Metasomal insertion on propodeum between hind coxae. Metasoma elongate, anteriorly cylindrical becoming dorso-laterally compressed apically. Metasomal segment 1 with T1 narrow anteriorly, posteriorly evenly broadened, with spiracles in front of midpoint; glymmae present (Rhyssa) or absent; T1 and S1 fused except in Rhyssa; sclerotized anterior section of S1 short and not reaching beyond midpoint. T2-5 generally rigid and strongly sclerotized, without distinct lateromedian rounded swellings or other surface irregularities. T8+9 of female elongate, apex modified as a horn or boss. S2-4 with pair of tubercles. Hypopygium of female short, transverse, folded on midline. Ovipositor straight or apically slightly decurved, projecting beyond apex of metasoma by a distance equal to or greater than 5x length of hind tibia; upper valve without dorsal subapical notch; apex of lower valve with conspicuous teeth.
The genera of Rhyssinae present in the Nearctic Region
- Epirhyssa Cresson, 1865; worldwide; one described Nearctic sp. (mexicana Cresson) (HOD)
- Megarhyssa Ashmead, 1858; Ethiopian, Holarctic, Oriental; four described Nearctic spp. (HOD)
- Rhyssa Gravenhorst, 1829; Holarctic, Oriental, Neotropic; seven described Neartic spp. (HOD)
- Rhyssella Rohwer, 1920; Holarctic; three described Nearctic spp. (HOD)
Identification and resources
Rhyssines are amongst the easiest of ichneumonids to recognize on account of the prominent transverse rugae on the mesoscutum and the horn-like projection on the end of the metasoma of females. The only other ichneumonid with similar mesoscutal sculpture in North America is Pseudorhyssa in the Poemeniinae, which has impressed oblique grooves anteriorly on tergite II (none are present in rhyssines) and dorsal carinae on the propodeum (no dorsal carinae are present in Rhyssinae). The tropical labenine genus Apechoneura also has mesoscutal rugae, but species of this genus have a complete areolet in the fore wing (the only tropical rhyssine Epirhyssa lacks an areolet). Apechoneura species also have the metasomal insertion far above the metacoxal insertions, and possess some propodeal carinae.
Key to genera of Nearctic Rhyssinae
· Downloadable pdf file of the key is available here
· Downloadable pdf file of the key figures is available here
Hymenoptera Online Database (HOD)
Keys to the Nearctic species are given by Townes & Townes (1960).